شنبه, ۸ اردیبهشت, ۱۴۰۳ / 27 April, 2024
مجله ویستا
ایو باکتری ها یا باکتری های حلالی بدون جدار سلولی
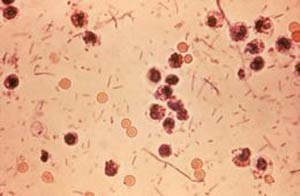
● مقدمه:
میکرواروگانیزم های مورد مطالعه در این تحقیق شامل دو گروه باکتری ها ی حلالی به نام ملیکیوتس که یکی از گروه گونه های پست ، سیتوزین مثبت و گرم مثبت که فاقد دیواره سلولی کلاسیک Mollicutes بوده ودیگری شامل دو باکتری قرمز رنگ بنام پروتیباکتری ها از گروه گونه های گرم منفی می بــاشد . بعضی ازاین ار گانیزم ها به آوندهای عآلی گیاهان حمله کرده و تولید بیماری می نمایند. بنابراین آنها جزء عوامل بیماریزای گیاهی محسوب شده در حالی كه انواع دیگر فقط بر روی بافتهای گیاهی مخصوصاً گلها مشاهده شده و بیماریزا نیستند. از میان باکتریهای بیماریزا بر روی آوندهای عآلی فقط نوع اسپیرپلاسماها قابل کشت می باشند در حالی که نوع فایتوپلاسماها و دو نوع از پرتی یو باکتریها هرگز کشت نشده اند. از زمان اولین گردهمایی مربوط به آلودگی باکترییایی ویا شبكه باكتری در محیطهای کشت بافت در سال ۱۹۸۷ که در آن گردهمایی ملیکیوتس های گیاهی برای اولین بار معرفی شدند تا به حال مطالعات زیادی در خصوص آنها صورت گرفته مخصوصاً در خصوص باكتریهای دارای دیواره سلولی که به آوندهای گیاهان حمله می کنند. پیشرفتهای مربوط به رده بندی ، خصوصیات طبقه بندی ، شناسایی و تشخیص این ارگانیزم ها دراین تحقیق بررسی گردیده كه غالبِأ بخاطر وجود این تکنولوژیهای جدید ویا اصلاح شده درزمینهی بیوشیمی مولکولی ، بیولوژی وژنتیک مخصوصاً در رابطه با زنجیرههای مولكول DNA ریبوزمی S۱۶ و مقایسه الكتروفریسیس به منظور اندازه گیری وآزمایش UGA با كدهای قابل استفاد ه بوده است. آزمایش UGA به منظورمتوقف كردن كدها ویا برای كد تریپتوفان، تعیین كلسترول ۸۰ مورد نیاز ملیكیوتس آزمایش، مربوط به سیستم قند تراسفرز (PTA ) احیا كننده و بالاخره برای احیاء فراگمنت های DNA مخصوص که دارای زنجیره ی احیاء پلی مرز ( (PCR جهت واکنشهای زنجیره ای برای شناسایی خصوصیات عوامل باکتریزا وعلل دیگرمورد استفاده قرار گرفته است. اثبات و بررسی مراکز و محل اولیه باکتریهای حلالی ازنوع ملیکیوتس: مقایسه زنــــــجیره های DNA ریبوزمی ۱۶S مربوط به گروه ملیكیوتس ها با باکتریهای دیگر نشان داده است که، ملیکیوتسها دارای یک شبکهی پلی لوژنتیک درخت مانند از نوع باکتریهای حلالی گرم مثبت می باشند . در حال حــاضر ملیکیوتس ها به علت تکامل بعضی ژنها ی خود را ازدست داده واندازه آنها تحلیل رفته است. در صورتی که نژاد اولیهی آنها باکتریهای گرم مثبت بوده که دارای (G+C)كمتری در ژن آنها وجــود داشته است. اقوام نزدیک باکتریهای حلالی که دارای دیوارهی سلولی بسته میباشند، شامل دو گونه گرم مثبت با G+C)) كم بنامهای clostridium ramosum و innocuum clostridium می باشند. این نوع کلستریدیاها شبیه ملیکیوتسهای اولیه بوده که انروپلاسماز واستروپلاسمازآنها هنوز از نــــــوع مخصوص انیربز می باشد که نشان دهندهی انیربیوسز وراثتی موجود در نژادهای اولیه آنها نیزهــــست؛ مقاومت موجود در ملیكیوتسها و همچنین در دو گروه کلستریدیال احتمالا از نژادهای اولیه آنها كه دارای(G+C) كمی در ملكول DNA بودهاند بهارث رسیده است.
● فایــــــــــلوژنتیک درختی ملیکیوتسها: این درخت پنج گروه فایلوژنتیکی را نشان می دهد
۱) گـــــــروه اسپیروپلاسما شامل دو گو نه جدید mesoplasma وEntomoplasma اما بعضی از انواع به خصوص Mycoplasma مثل M.mycoides نیز مشاهده میگردد.
۲) گـــــــروهmycoplasma pneumonias .
۳)گــــــروه mycoplasma hominis))
۴) گروه انیرو پلاسما اكولید پلاسما كه در این گروه فایتو پلاسما خوشهای می باشد.
۵) گروه استرو پلاسما كه فقط دارای یك نوع باكتری می باشد . ملیكیوتس های كه دارای شاخه های فایلوژنتیكی نزدیك به هم با (G+C) كم وگرم مثبت بوده در آنها از دست رفتن دیواره سلولی درهمان اوایل صورت گرفته است. (شكل۲) كه علت آن از دست رفتن ژن (شكل۲ GL ) در طول تكامل بوده است . شاخه شاخه شدن گروه استرولی پلاسما نشان دهنده ی ازدست دادن دیوارهی سلولی در مراحل اولیه میباشد . احتمالاً این مطلب صحیح نمیباشد، زیرا عمل شاخه شاخه شدن در این گروه هنوز صورت نگرفته است؛ در كد ژنتیكی یونیورسال فقط یك كد برای تریپتفان وجود دارد كه عبارتند از ۳ .۵uGG ملیكیوتسها به عنوان ارگانیزم های دارای ( (G+Cكم موفق به ساخت كد جدید تریپتفان با G كمتر بنام ۵UGA شده اند كه در آن ۳A بجای ۳G در UGG قرار گرفته است. این مطلب به صورت Trp =UGA نشان داده شده است. برای انجام این جابجایی آنها یك مولكول RNA جدید را با آنتی كد ۵VCA۳ كه قابلیت ترجمه UGA و UGG را دارد بوجود آوردهاند. ((۴ را ببینید )) بنابراین در تكامل اخیر ملیكیوتسها مثل اسپیروپلاسما، انتموپلاسما، مزوپلاسما، مایكوپلاسما و یوریوپلاسما ژن UGA به عنوان كد تریپتوفان مورد استفاده قرار گرفته است در حالی كه در ملیكیوتسهای اولیهی مثل انیرپلاسما، استروپلاسما، اكولی پلاسما، فایتوپلاسما ژن UGA به عنوان كد متوقف كننده مورد استفاده قرار گرفته است؛ هر چند كشت فایتوپلاسما امكان پذیر نمی باشد اما گفته می شود كه شاخههای آنها به اكلیپلاسما شباهت داشته وآنها ملیكوتسهای حقیقی میباشند.
● رده بندی ملیكیوتسها تا حد گونه : تعریف گونه های ملیكیوتس ها بر اساس خصوصیات مختلفی مثل انیریوسیز، هلیكال مرفولوژی، احتیاجات كلسترول ویا توین ۸۰، درجه حرارت مناسب رشد وهیدرولیز اوره صورت می گیرد. اندازهی ژنوم به عنوان یك خصوصیت مهم در تعریف گونه مد نظر قرار گرفته است با بوجود آوردن الكتروفرسیس ضربانی(PFGE ) كه یك تكنولوژی دقیقتر وبهتری نسبت به روش اولیه احیا جنبشی میباشد نشان داده شده است كه در میان گونههای زیاد اندازهی ژنوم بسیار متفاوت میباشد؛ نیاز كلسترول برای مدت زیادی به عنوان یك عامل اصلی در تشكیل رده بندی در بین ملیكیوتسها اهمیت داشت. اما كشفیات جدید این موضوع را در طبقه بندی ملیكیوتسها كم اهمیت كردهاست. گونههای زیادی از باكتریهای حلقوی (اسپیروپلاسما ) مثل S.apils S.floricola S.chinense S.diabroticae بدون حضور كلسترول نیز رشد ونمو كردهاند این نشان دهندهی این مطلب است كه خانوادهی اسپیروپلاسماتسی از نظر ژنتیكی به دو شاخه تقسیم شده اند ویك گونه جدید بنام هلیكوپلاسما كه شبیه Mollicutes حلالی بوده وبرای رشد احتیاجی به استرول ندارد نیز بوجود آمده است؛ همچنین ممكن است نیاز كلسترول به عنوان یك عامل مهم در طبقه بندی Mollicutes كاملاً از بین رفته باشد.
باالاخره خانوادهی انتمپلاسماتالیس كه شامل گونه های انتمپلاسما ، مزوپلاسما و اسپایروپلاسما میباشد، براساس محل زندگی رده بندی شده است . در حقیقت بیشترین میزبان این خانواده حشرات می باشند و عامل انتقال دهندهی آنها زنجیرههایی می باشند كه بر روی برگ گیاهان زندگی می كنند.
روشهای تشخیص مولكول در خصوص مایكوپلاسمولوژی و مطالعات آنها اخیراً قابل دسترس می باشد. باكتریهای دارای دیوارهی سلولی عامل بیماری در سلول آوندی لیبروباكتری وفلوموباكتری (پرنی یو باكتری ها ) Candidatus phlomobacter fragariae نامگذاری گردید.
● پروتی باكتریهایی از نوع (گاما) به سه زیرگروه مهم تقسیم می شوند
۱) باكتری های فتوسنتز كننده ازنوع سولتور قرمز مثل chroatiun
۲) گونه هایی كه با بیماریهای لكهای همراه هستند.مثل Legionella
۳) مخلوطی از انواع غیرفتوسنتز كننده مثل Endosymloiontsو pseudomonadsو Escherichia coli كه به مورچه ها حمله میكنند. Buchaera كه به شته و مگس تسه تسه ومگس سفید و زنجره حمله میكند كه گونهBacter phlomo در همین زیر گروه كه بیشترین انگلهای باكتری حشرات در آن موجود می باشند یافت میگردد . بر اساس خاصیت توالی در مولكول DNA ریبوزمی ۱۶S موجود در باكتریBacter phlomo در گیاه توت فرنگی یك نوع pcr جدید بوجود آمده است ؛كه برای اولین بار شناخت عامل بیماری توت فرنگی را در گیاهان آلوده امكان پذیر مینماید. بدون شك برای شناخت حشره ناقل بیماری نیز مناسب می باشد. به طور خلاصه هرچند كه كه عوامل زنده ومیكروارگانیزم های موجود درسلول های آوندی معمولاً همراه با بیماری سبز مركبات و زردی لبه ی برگهای توت فرنگی می باشند، كشت آنها در محیط آزمایشگاهی امكان پذیرنیست. اما از طریق بدست آوردن مولكول های DNA ریبوزمی ۱۶S روش pcr متوالی ومقایسه آنها با DNA ریبوزمی۱۶S موجودات دیگرمی توان آنها را شناسایی كرد. این تحقیق نشان می دهد كه شبه باكتریهای اولیه ، شبه باكتری های حقیقی هستند كه دقیقاً به آنها پرتو باكتری گفته میشود. لیبروباكتری های سبز دارای یك زنجیره در زیر مجمموعه مربوط به پرتوباكتریها میباشند اما باكتریphlombacter در توت فرنگی یك زنجیره جدیدی است كه مربوط به پرتوباكتری نوع گاما( ) می باشد. هر دو زیر مجموعه آلفا و گاما دارای باكتری هایی می باشند كه به حشرات نیز حمله می كند. بنابراین انتقال لیبرو باكتری بوسیله حشرات یك امر معمولی بوده و عامل انتقال باكتریplomobacter نیزمربوط به گروهای شبه باكتری eubacteria نیزمی باشد.عوامل بیماری محدود در سلول آوندی باكتریهای بدون جدار ،اسپیروپلاسما و فایتوپلاسماست، برعكس شبه باكتری ها (eubacteria ) كه در گذشته مورد بحث قرار گرفتند، اسپیروپلاسماها وازنوع Mollicutes بوده كه فاقد دیواره سلولی می دهد.
اسپیروپلاسماها حلالی شكل بوده ودر محیط كشت قابل كشت میباشد. اما فایتوپلاسماها حلالی شكل نبوده و هر گز قابل كشت نمیباشند. اسپیروپلاسماها شامل گروهای فائلوژنتیك با كد UGA مربوط به تریپتوفان بوده اما فایتوپلاسماها جزئی از گروه اكولی پلاسما بوده كه كدUGA آنها متوقف كننده می باشد. همچنین خصوصیات فایتوپاتوژنتیكی یا بیماریزائی در اسپیروپلاسما وهمه فایتوپلاسماها درمحلهای یكسان در گیاهان یافت میشود. این محلها معمولاً سلولهای آوندی مَشبك در بافتهای آوندی میباشد. اسپیروپلاسماها و فایتوپلاسماها دارای دو میزبان یعنی گیاهی و حشره مخصوصاً زنجره ها هستند.● اسپیروپلاسماها : اسپیرو پلاسماها در طول مطالعات مربوط به دو بیماری گیاهی كشف گردیدند،این دو بیماری عبارتند ازبیماری كوتولگی ذرت و بیماری خشكیدگی مركبات ، عامل خشكیدگی اولین Mollicutes است كه منبع آن گیاه می باشد. بوسیله كشت در سال۱۹۷۰بدست آمد. این ملیكیوتسها دارای ارگانیزمهای بدون دیواره سلولی بوده وساختمان آنها حلالی شكل میباشد. عامل خشكیدگی در سال ۱۹۷۳ بنام spiroplasma citri شناخته شد و اولین اسپیروپلاسمائی بود كه در آزمایشگاه كشت و مشخصات آن توسط همكاری های بین المللی ارائه گردید. عامل بیماری كوتولگی ذرت در سال۱۹۷۵در آزمایشگاه كشت شد و مشخصات كامل آن درسال۱۹۸۶ ارائه وبنام spiroplasma kuukelliiنامگذاری گردید. سومین نوع اسپیروپلاسمایی گیاهی مربوط به سلول آوندی مشبك بنام spiroplasma phoeniceum در سال ۱۹۸۳و۱۹۸۴از گیاه تلفونی آلوده درسوریه جداسازی وكشت گردید.میزبانهای مختلف این سه نوع اسپیروپلاسمای بیماریزا قبلاً مورد مطالعه قرار گرفته بود. از زمان تحقیقات اولیه بر روی s.kunkellii , s.citri تا به حال تعداد زیادی اسپیروپلاسما شناخته شده كه ناقل آنها زنجره بوده كه از طریق آن اسپیروپلاسما از گیاه آلوده به گیاه سالم منتقل می گردد۰به خاطر قابل كشت بودن اسپیروپلاسما مطالعات زیادی بر روی آنها از سال۱۹۷۰تا به حال صورت گرفته است. بعد از ۲۵ سال تحقیق بر روی خشكیدگی مركباتspiroplasma citri به این نتیجه رسیدهاند كه احتمالأ باكتری spiroplasma citri مهمترین باكتری شناخته شده از این نوع بوده و برای تجزیه های ژنتیكی مناسب می باشد.
● فیتوپلاسماها : آقای ده ئی و همكارانش مشاهده كردند كه در سلول های آوندی مشبك مربوط به گیاهان آلوده به بیماری زردی ،عوامل بیماری یا میكروارگانیزمها از نظر ساختمانی شبیه به مایكوپلاسماهای حیوانی می باشد ، بر اساس این شباهت عوامل شبه مایكوپلاسما(mlo ) نامیده شدند. امروز بیشتر از ۳۰۰ گونه گیاهی مختلف متعلق به ۹۸ خانواده كه با شبه مایكوپلاسماها آلوده می باشند ، شناسائی شده است. با وجود كوششهای بسیار زیاد ، شبه مایكوپلاسماها(mlo )هرگز قابل كشت در آزمایشگاه نمیباشند. ماهیت اصلی آنها تا سالهای زیادی قابل شناسائی نمی باشد. فقط وقتی كه تكنیكهای بیولوژی مولكولی در خصوص شبه مایكوپلاسماها (mlo ) بكاربرده شد ، مشخصات و خصوصیات آنها به سرعت شناخته شده و مورد برسی قرا گرفت.
● امروز میدانیم كه شبه مایكوپلاسماها درحقیقت عضوی ازكلاس ملیكیوتس ها می باشند زیرا:
۱) ماده( G+C ) موجود درDNA آنها به میزان ۲۵-۳۰ درصدشبه میكیوتسها قابل كشت می باشد
۲) اندازه ژنم آنها با استفاده از روش الكتروفورسز ضربانی كوچك بوده و به اندازه ۶۰۰ تا ۱۲۴۰ kbp می باشد كه به گونه های میلیكیوتس مربوط میشود.
۳) DNA استخراج شده از برگهای آلوده به شبه مایكوپلاسما(mlo ) برای تعیین DNAریبوزمی ۱۶Sبا استفاده ازخصوصیات این نوع DNA درشبه باكتریها مورد استفاده قرارگرفت مولكول DNA ریبوزمی ۱۶s در شبه باكتری ها(mlo )می تواند متوالی ورشته ایی باشد. این تحقیق بر روی انواع زیادی از شبه باكتری ها(mlo ) صورت گرفت. مقایسه توالی نشان می دهد شبه مایكوپلاسماها (mlo ) باید ازنظرژنتیکی گیاهی به باکتری های گروه اكولی پلاسما ویا انیروپلاسما باشند .
۴) روابط وراثتی آنها گروه اكولی پلاسما به خاطر وجود کد UGA به عنوان کد متوقف کننده نه به عنوان کد تریپوفان مورد تائید قرار گرفت . این نشان می دهد که شبه مایکو پلاسماها( MLO ) مثل اکولی پلاسما از نظر فایلوژنتیکی یا ژنتیک گیاهی به ملیکیوتسهای اولیه شبیه بوده واین مطلب متضاد شباهت آنها به اسپیروپلاسماها می باشد . مدل اولیه و مخصوص DNA ریبوزمی ۱۶Sمربوط به فایتو پلاسما طراحی گردیده است. DNA ریبوزمی مربوط به هر باکتری از نوع فایتو پلاسما می تواند به صورت رشته متوالی باشد و برای مطالعات ژنتیکی- گیاهی یا فایتوژنتیک استفاده گردد . همچنین می تواند برای آنالیزپلیمرفوس ( RFLP ) نیز به کار رود . کمیته رده بندی ملیکیوتس ها در سال ۱۹۴۴ شبه مایکو پلاسماها را بعنوان عضوی از کلاس ملیکیوتس دانست و واژه فایتو پلاسما را به جای شبه مایکوپلاسماMLO)) قرار داد. این كمیته همراه با تیم تحقیقی فایتوپلاسما بنام IRPCM پیشنهاد كردند كه واژه Candidatus برای گروهای اصلی فایتوپلاسما مورد استفاده قرار گیرد . هر گروه بیان کننده گونه های مشخص فایتو پلاسما می باشد . تا کنون ۱۴ گروه از طریق آنالیز DNA ریبوزمی ۱۶S بدست آمده است. فایتوپلاسما یی که درلیموایجاد بیماری میکند اولین فایتوپلاسمایی است که به عنوان Candidatus مورد بررسی قرارگرفته است. واسم آن عبارتنداز Candidatus phytoplasma aurantifolia. بررسی این فایتوپلاسما بر اساس مولکول DNA ریبوزمی ۱۶S متوالی، محل توالی ۲۳S,۱۶S اندازه ژنم (genome) هیبریدازیسیون و تشابهات ژنتیکی با گروههای دیگر فایتوپلاسماها صورت گرفته است. رفتار فایتو پلاسما در محیط کشت گیاهی نیز بسیار جالب میباشد . فایتوپلاسماهای موجود در درخت سیب می تواند دربافتهای زایشی گیاه به طورطبیعی مستقر باشند اما همچنین فایتوپلاسماها دریافتهای آوندی ایجاد بیماری می کنند. مطالعات نشان می دهند که، حضور فایتوپلاسما در درخت سیب میتواند حتی بعد از ۱۰ سال ایجاد بیماری نمایید. آنالیزRFLP در خصوص DNA کرومززومی نشان داد که هیچ گونه تفاوتی بین فایتوپلاسماها موجود در درخت سیب نمیباشد . پارامترهای رشد و غلظت فایتوپلاسما به منظور مقایسه رفتار چهار نمونه جدا شده از درخت سیب وهمچنین درشرایط مختلف مورد مطالعه قرار گرفت . مقایسه آمار بدست آمده بعد از ۲ الی ۸ سال نشان دهنده هیچ گونه اختلاف فاحشی نمیباشد.نگهداری فایتوپلاسمایی روی میزبانان درمحیط آزمایشگاه به مدت ۸ سال و سپس انتقال آنها به گیاه اصلی دوباره تولید بیماری کرده وعلائم مشابهی را مثل کوچک شدن برگها و بزرگ شدن دمبرگها وکم شدن رشد ایجاد می نمایید. بنابراین ایزوله کردن فایتوپلاسمای سیب درشرایط آزمایشگاه به مدت ۸ سال هیچگونه تغییری در آنها ایجاد نمی نماید. به مدت ۸ سال بر روی میزبان غیر طبیعی pyronia veitchii نگهداری می شد، هیچگونه علائم بیماری بر روی گیاه میزبان مشاهده نگردید هر چند در تمام گیاهان مورد آزمایش حضور داشت . اما درشرایط آزمایشگاهی فایتوپلاسما برروی گیاهانی مثل گل داوودی، گلایول، شمعدانی گیاه گل تلفونی و بادمجان ایجاد بیماری مینماید. مطالعاتی که بامیکروسکوپ الکترونی صورت گرفته است نشان دهنده مقدار زیادی فایتوپلاسما در محل سلولهای آوندی مشبک می باشد . کنترل بیماری در شرایط مزرعه با استفاده از تتراسایکلین صورت می گیرد در صورتی که مقدار آن در محیط کشت جهت کنترل به مقدار(۱۲.۵mgml-۱ ) می باشد . اگر گیاه فقط یک بار مورد مطالعه قرار گیرد ، علائم بیماری باشدت بیشتری دوباره ظاهر می گردد.
بنابراین گیاه باید برای مدت زیاد و حتی ماهها به وسیله تتراسایکلین مورد درمان قرار گیرد . آنتی بوتیک های دیگر مثل اریترومایسن واسترپتومایسین نیزتا حدودی درمان کننده میباشند؛ اما به خوبی و موثری تتراسایکلین نیستند. گرما درمانی در درجه حرارت ۳۲ تا ۴۳ درجه سانتیگراد به مدت ۴ ماه اصلاً موثر نمیباشد.
این تحقیق بر روی انواع زیادی از شبه باكتری ها(mlo ) صورت گرفت در خصوص تاثیر متقابل بین عوامل بیماری زا و گیاهان میزبان و همچنین حشرات ناقل ،مطالعات اندكی صورت گرفته است. مطالعات انجام شده بر روی بیماری spiroplasma citri نشان دهندهی اهمیت تأ ثیرمتقابل و زمان بررسی آن می باشد باكتری S. Citri یك عامل بیماری گیاهی است میزبان آزمایشگاهی این بیماری گیاه گل تلفونی بنام catharantus roseus است در طبیعت آلودگی گیاهان فقط بوسیله حشرات ناقل بیماری صورت میگیرد زنجیره ای بنام circulifer haematoceps مهمترین ناقل این بیماری دركشورهای منطقه مدیترانه و غرب آسیای می باشد. بنا براین بیماری s. citri دارای دو میزبان بوده كه در بدن آنها تولید مثل مینماید. یكی زنجیره و دیگری گیاه. در اینجا ما به ژنهائی كه در بین اسپیروپلاسما ودو میزبان آن تاثیر متقابل ایجاد مینمایند علاقه مند میباشیم. این ژن ها بوسیله جهش و روشهای نمایشی ( screenig) قابل ردیابی و شناخت می باشند.
یك روش جدید برای جهشهای ژنی بوسیله تزریق پرتوهای رادیم tn ۴۰۰۱ به درون ژن های باكتری s. citri بوجود آمده است. این روش برای ساختن ژنهای عامل انتقال باكتری و مطالعهی آنها مورد استفاده قرار می گیرد. اولین عامل انتقال مورد استفاده فرم متشابه RF باكتری S. citri . به صورت ویروس spv۱ بوده اما عامل انتقال RF پایدار نبوده و DNA تزریق شده به سرعت تحلیل می رفت. این پدیده بوجود آمدن تركیبات همولوگوس RF شد و حذف مولكول دخالت داشت هر چند پروتئین reca جهت تشكیلHR كه در تشكیل همه گونه باكتری s. citriبسیارناچیز و كم بود. راه دوم استفاده از مركز تولید و منبع به وجود آمدن باكتری s. citri و DNA آن بود. (oric)تا بتوان مقدار زیادی پلاسمید مصنوعی تولید كرد. این پلاسمیدها با موفقیت به عنوان عامل ژنتیكی مورد استفاده قرار گرفتهاند. آنهایی كه دارای رشتههای coLEL بودند دربین باكتریهای E.coliوs.citri به عنوان حائل فعالیت میكردند. بعضی ازاین پلاسمیدها به صورت كرومزومهای خارجی رفتار كرده و بعضی دیگر به ژنهای اسپیروپلاسما در منطقه ی oric ملحق میشدند.همراه با این پلاسمیدها مادهی اسپیرالین موجود درباكتری S. phoeniceum به مقدار زیاد در باكتریs. citri متوان تزریق كرد همچنین در زیر گونههای باكتری s.citri و ژن های آن میتوان به آسانی به زیر گونههای دیگر انتقال داد. در این خصوص ژن g۱۱۳ از كپی زیر گونه ها جهت جهش ژنتیكی tn ۴ool انتخاب گردید. زیرا این ژن میتواند به آسانی توسط زنجره به گیاه گل تلفونی انتقال یابد. در حدود ۱۰۰۰ نوع ماده جهش دهنده tn ۴ool از باكتری s.citri بدست آمده است ماده ی جهش دهنده شماره۵۵۳ در بدن حشره به خوبی رشد میكند و به آسانی به اندام گیاه گل تلفونی انتقال یافته و به مقدار زیاد در آن تولید می گردد اما تا وقتی كه عكس العمل صورت نگرفته است، هیچگونه علائمی ایجاد نشده و نوع وحشی اسپیروپلاسما به صورت طبیعی باقی می ماند. ماده جهش دهنده شماره ۴۷۰ در بدن زنجیره ازدیاد و تولید نشده و در نتیجه در بدن گیاه انتقال نمی یابد . ژنهای جهش دهنده كه در آنها خاصیت انتقال وجود دارد شناسایی شده اند در مادهی جهش دهنده شماره ۵۵۳ ژن موثر و تاثیر گذار در محل قند فروكتوز قرار دارد و اصلا ایجاد بیماری نمی نماید. قند فروكتوز قابل انتقال نبوده ونمی تواند وارد فعالیتهای سوخت و ساز در سلولهای اسپیروپلاسما گردد. حال چطور حضور قند فروكتوز باعث بوجود نیامدن علائم بیماری می گردد معلوم ومشخص نبود وباید در این خصوص تحقیق كرد. امید است كه این مطالعات نه تنها به فهمیدن وشناخت اثر متقابل بین میزبان وانگل كمك كند بلكه ارائه دهندهی راههای جدیدی برای كنترل بیماری در گیاهان می باشد .
عبداله کرمی
مطالعه وتحقیق توسط آقایان جی ام بوهو منیک گارنیر ازکشورفرانسه
مطالعه وتحقیق توسط آقایان جی ام بوهو منیک گارنیر ازکشورفرانسه
منبع : بزرگترین وب سایت فارسی گیاهپزشکی



همچنین مشاهده کنید
















































































































نمایندگی زیمنس ایران فروش PLC S71200/300/400/1500 | درایو …
دریافت خدمات پرستاری در منزل
pameranian.com
پیچ و مهره پارس سهند
خرید میز و صندلی اداری
خرید بلیط هواپیما
گیت کنترل تردد
حجاب دولت چین ایران رئیس جمهور پاکستان رئیسی گشت ارشاد دولت سیزدهم کارگران رهبر انقلاب سریلانکا
کنکور سیل سردار رادان تهران قم سازمان سنجش فضای مجازی اصفهان سلامت شهرداری تهران پلیس زنان
خودرو دلار واردات خودرو قیمت خودرو آفریقا تورم قیمت دلار قیمت طلا بازار خودرو سایپا ایران خودرو ارز
پایتخت سریال پایتخت خانواده تلویزیون موسیقی ترانه علیدوستی فیلم سریال سینمای ایران مهران مدیری کتاب
کنکور ۱۴۰۳ عبدالرسول پورعباس
فلسطین غزه اسرائیل رژیم صهیونیستی آمریکا جنگ غزه روسیه حماس طالبان اوکراین ایالات متحده آمریکا ترکیه
پرسپولیس فوتبال آلومینیوم اراک جام حذفی استقلال فوتسال بازی بارسلونا تراکتور باشگاه پرسپولیس باشگاه استقلال لیگ برتر انگلیس
همراه اول ناسا الماس تسلا فیلترینگ سامسونگ
دندانپزشکی مالاریا آلزایمر پیری کاهش وزن سلامت روان زوال عقل