جمعه, ۱۷ اسفند, ۱۴۰۳ / 7 March, 2025
مجله ویستا
پاسخهای سلولی به محیط نانوالیافی
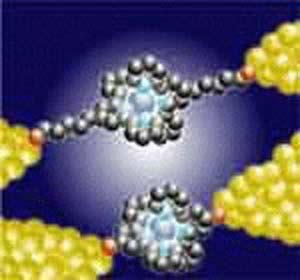
سلولها در محیط زنده بدن توسط یک ماتریکس شبکهای برونسلولی و سه بعدی از نانوالیاف ۱(ECM) دارای لیگاندهای شیمیایی متفاوت که با پذیرندههای سطح سلول برهمکنش میکنند، نگهداری میشوند [۴-۱]. آنچه باعث ارتباط ECM و سلول میشود فرایند دینامیکی و پیچیدهای است که طی آن خواص شیمیایی و فیزیکی ECM، موجب پاسخهای سلولی متفاوتی میشوند [۵ و۱]. بنابر این برای آنکه یک بستر مهندسی شده دقیقاً همان کار ECM طبیعی را انجام دهد، باید همان (نقشه) نانوالیافی و سه بعدی بودن و نیز فراوانی موتیفهای شیمیایی آن را دربرداشته باشد [۷و۶]. پیشرفت فناورینانو در سالهای اخیر امکان طراحی و مهندسی مواد زیستی جدید با این سطح از پیچیدگی را فراهم کرده است [۹-۷].
استفاده از این مواد زیستی پیچیده برای کاربردهای خاص توسط مصرف کننده نهایی، مستلزم انجام یک فرایند محاسباتی بازگشتی برای درک مکانیسم هدایت کننده برهمکنشهای سلول- شبکه میباشد، تا به این ترتیب بتوان خواص ماده زیستی را کنترل کرد و در نهایت به پاسخهای سلولی مناسب و دلخواه دست یافت . در اینجا ما بر اهمیت ویژگیهای نانوالیاف سه بعدی محیط برونسلولی و نقش آن در تنظیم پاسخهای سلولی دارای تفکیک موضعی یا زیرسلولی به روشی وابسته به زمان- مکان تأکید میکنیم. همچنین فناوریهای نوین ساخت و تعیین مشخصات محیطهای نانوالیافی در کاربردهای مرتبط با آن، از دیگر مواردی است که در این مقاله بیشتر به آن توجه میشود.
● تأثیر محیط نانوالیافی بر پیامرسانی و فنوتیپ۱ سلول
سلولها از طریق پروتئینهای سطحی خود از قبیل اینتگرینها۲ با محیط نانوالیافی خارجی پیرامون خود برهمکنش میکنند و در اثر این برهمکنشها، مسیرهای مختلف سیگنالدهی که فرایندهای سلولی از قبیل شکل سلول، تحرکپذیری و تكثیر سلولی را تنظیم میکند، فعال میشوند (شکل ۲). آگاهی از این پاسخهای (زیستی) ویژه، که با جنبههای مختلفی از محیط نانوالیافی تحریک میشوند- در هدایت طرحها و مهندسی بسترهای جدید که به تقلید از برهمکنشهای سلول- ماتریکس داخل بدن، ساخته میشود اهمیت بسزایی دارد.
● تأثیر توپوگرافی سه بعدی
برخلاف بسترهای دوبعدی پهن و صلب، این نانوالیاف سه بعدی امکان برهمکنش سلولها با یک حلقه سهبعدی انعطافپذیر را فراهم میکند. این نانوالیاف سه بعدی همچنین باعث ایجاد فتوتیپهای سلولی مشابه فتوتیپهای درون بدن میشود و سلولی شکلزایی بافت را نیز افزایش میدهند [۱۳-۱۰]. رویدادهای مولکولی مربوط به برهمکنش سلولها با این نانوالیاف سهبعدی نیز متفاوت از موارد مشابه در بسترهای دوبعدی از قبیل ظرفهای کوچک مصنوعی کشت میکروب میباشد. سه بعدی بودن این نانوالیاف به تنهایی در فعال کردن مسیر تبدیل سیگنال با واسطه Rac۳ داخل فیبرو بلاستهایNIH۳T۳ و سلولهای عادی کلیه موش، نقش دارد، به خصوص اگر روی نانوالیاف الکتروریسی۴ شده و در غیاب ماکرومولکولهای ECM کشت شوند [۱۴].
حرکت فیبروپلاست در شبکههای کلوژنی سه بعدی (که در شکلگیری دوباره کلوژن طی روند بهبودی زخم اهمیت دارد) ، تا حد بسیار زیادی به زنجیر سنگین میوزین غیرعضلانی Nanomuscle Myosine Heavy Chain II-B۱ بستگی دارد. این در حالی است که حرکت سلول روی سطوح دو بعدی صلب از این ویژگی برخوردار نیست [۱۵]. پاکسیلین (در چسبندگی موضعی) و اینتگرین (در چسبندگی رشتهای) ، برخلاف بستر دوبعدی با دو ناحیه چسبندگی مجزای موضعی و رشتهای کلاسیکی روی آن، حالت سه بعدی موضعی شده مشترکی پیدا میکنند. [۱۶]. اعمال فشار مکانیکی و پهن کردن مصنوعی ماتریکسهای نانوالیافی سه بعدی به منظور ایجاد شبکههای دو بعدی صاف مجازی، باعث از دست رفتن مشخصه موضعیسازی مشترک سهگانه اینتگرین، پاکسیلین و فیبرونکتین موجود در چسنبدگیهای شبکه سهبعدی میشود [۱۶]. لذا نقشهبرداری نانوالیافی سه بعدی، تأثیر بسیار زیادی بر برهمکنشهای سلول-ECM در سطح مولکولی خواهد داشت.
● تأثیر خواص مکانیکی
سلولها به لیگاندهای شیمیایی و نیز محرکهای مکانیکی متنوع محیط پیرامونی خارجی پاسخ میدهند [۱]. پخش کردن سلولها روی بسترهای صلب باعث تکثیر آنها میشود و lineage differenrition تمایز دودمان و سلولهای بنیادی (stemcell) را تغییر میدهد [۱۹]. سلولها به نیروهای حاصل از تنش برشی و فشردگی حساس بوده و در نتیجه آن مدلسازی مجدد و خواص مکانیکی ساختار بافت تحت تأثیر قرار خواهد گرفت [۲۳-۲۰]. در وضعیت سه بعدی، باز هم سلولها به صلب بودن نانوالیاف حساس هستند [۲۵ و ۲۴و ۱۶]. نیروهای خارجی هم از طریق مسیر ارتباطی ECM- اینتگرین- سیتواسکلتون۲ آرایش چسبندگی موضعی۳ که تیروزین کیناز فسفاتاز و كینازهایSRC SFKS را تحریک کرده و موجب فعالسازی پروتئینهای G کوچک و کینازهای MAP۴ و در نهایت تنظیم رویدادهای پایین دستی سلولی میشوند [۲۶]. باید توجه داشت که کمی کردن سیستماتیک این رویدادها و نیز ایجاد مدلهایی از پاسخ سلول به خواص مکانیکی و شیمیایی نانوالیاف به منظور هدایت روند توسعه محیطهای نانوالیافی سه بعدی به سمت مصرف و کاربرد، اهمیت دارد.
● تأثیر محدودیتهای زمانی- مکانی بر محرکهای برونسلولی
پاسخهای سلولی به خواص فیزیکی و شیمیایی نانوالیاف سه بعدی، غالباً به مکان و زمان بستگی دارد [۲۹-۲۷]. هنگامی که سلولهای بنیادی عصبی و سلولهای ماهیچهای صاف۵ به روش الکتروریسندگی کشت شوند، خود را در امتداد این الیاف کشیده و جهت میدهند [۳۱ و ۳۰]. حساسیت نانوالیاف به ترتیب مکانی، احتمالاً ناشی از برهمکنشهای موضعی سلولها با محرکهای برونسلولی است که در سطح تفکیک زیرسلولی روی میدهند. اغلب پروتئینهای ساختاری یا سیگنال دهندهای که در حسگری صلب، (تکیهگاه ماتریکسی برون سلولی) و مهاجرت سلولی حضور دارند، در نقاط خاصی از سلول از قبیل نوک جلویی فیبرو بلاستهای مهاجر یا نقاط مربوط به چسبندگی موضعی روی میدهد و در نقاط دیگر و حتی در فاصله ۵/۰ میکرومتر دورتر، چنین پدیدهای روی نمیدهد [۲۸].
یک اتصالECM- اینتگرین- سیتواسکلتون منفرد میتواند در نقطه معینی از سلول و در پاسخ به نیرویی خارجی از سلول و بدون تحت تأثیر قرار دادن اتصال مجاور ایجاد شود[ ۳۲ و۳۳]. خوشه شدن مولکولهای اینتکرین و تشکیل چسبندگی موضعی و الیاف فشاری ، تنها هنگامی روی میدهد که تعداد کافی از جایگاههایRGD، داخل ناحیه مجزایی با اندازه کوچکتر از ۷۰ نانومتر جمع شوند [۲۹-۲۷].
همچنین مولکولهای همراه با اتصالات اینتگرین- سیتواسکلتون موجود در چسبندگی سلول- ECM، به سخت بودن این نانوالیاف ECM حساس هستند [۳۸-۳۵و ۳۲] و طی چرخه زندگی نقاط چسبندگی، همراه با گذشت زمان، تغییر میکنند [۳۴]. شکلگیری تماسهای سلولی با ECM یک فرایند پیوسته نیست اما شامل چرخههای انقباض و رها شدن است. دانشمندان اخیراً نشان دادهاند که وجود نانوالیاف خارجی درهپاتوسیتها (سلولهای کبدی) در زمانها و توالیهای مختلف از کشت ساندویچی، پاسخهای متفاوتی را نشان میدهد [۳۹].
پس میتوان گفت سلولها نسبت به محدودیتهای زمانی و مکانی محرکهای محیط خارجی پیرامونی حساس هستند. به نظر میرسد مجموع پاسخهای موضعی سلول به این محرکهای برونسلولی طی زمان، بتواند به تعیین شکل و بیان ژنی سلول کمک نماید [۴۰ و ۲۶]. بنابراین مهندسی و درک پاسخهای موضعی سلولی به توزیع زمانی و مکانی محرکهای برون سلولی، در توسعه بیشتر محیطهای نانوالیافی سه بعدی پیرامونی اهمیت بسیاری دارد.
● تأثیر محرکهای برونسلولی بر پاسخهای موضعی سلولی
با توجه به تحریک حاصل از محرکهای برون سلولی (جدا از هم) و به حد کافی قوی، سلولها در ابتدا به طور موضعی و سپس همگی به این محرکها پاسخ میدهند. شبكه آندوپلاسمی ER۱ و بسیاری دیگر از ترکیبات مرتبط با شبكه آندوپلاسمی، معمولاً به صورت یکنواخت و با ساختاری شبکه مانند داخل سیستوپلاسم سلول و در یك محیط كشت دو بعدی توزیع میشوند . این ترکیبات در صورت قرار گرفتن در معرض تحریکات خارج از سلول، تماس سلولی، تکثیر و یا مهاجرت، میتوانند به صورت پویا آرایشی دوباره پیدا كنند.
هنگامی که دانههای پوشش شده با فیبرزنكتینبا سطح بالایی سلول برهمکنش میکند، برخی از پروتئینهای غشایی ER از قبیل کینکتین، پروتئین همراه با گیرنده(RAP) ، کالرتیکولین۱ [۴۱] و فاكتور طویل كننده یوركاریوتی((EEF-۱&#۹۴۶; ( EEF) به سرعت به محل ترکیبات اینتگریندار AC که به صورت موقت در اطراف بید پوشش شده با فیبرونكتین شکل گرفتهاند منتقل میشوند. این ترکیبات به فیبرونکتین پخش شده در محلول یا فیبرونکتینی که به طور یکنواخت روی یک سطح دوبعدی بزرگ پخش شده باشد، پاسخ نمیدهند (شکل ۴). پژوهش دانشمندان نشان داده شده است که EEF-۱&#۹۴۶; از طریق کینکتین باعث نگه داشتن کل ترکیب EEF روی غشاء ER شده و سنتز پروتئینهای غشاء/ ترشحی۲ و سیتوزولیک۳ را تنظیم میکند [۴۳].
به علاوه محققان مشاهده کردند که ریبوزومها و MRNA، IACهای اطراف دانههای پوشیده شده از فیبرونکتین را هدف گرفته و قبل از آنکه تغییرات نسخهبرداری را بتوان ردیابی نمود، سنتز پروتئینی سریعی روی میدهد [۴۵و ۴۱]. بنابر این میتوان سنتز موضعی پروتئینها را یک فرایند سلولی کلی فرض نمودکه در پاسخ به نشانهها و محرکهای قوی، گسسته و موضعی شده خارج از سلول به روشی وابسته به مکان و احتمالاً زمان روی میدهد (شکل ۵). بر اساس این فرضیه ترجمه محلی۴ (LTH)، کارخانه سنتز پروتئین یک سلول- شامل ریبوزومها، MRNA و تنظیم کنندههای انتقال (از قبیل میکرو RNA) به محلی نزدیک محرک خارجی – که میتواند دانههای پوشیده شده با فیبرونکتین [۴۱]، نیروی وارد شده توسط انبرکهای نوری [۴۶] یا دیگر نقاط اتصال زیر لایه صلب باشند- حرکت میکند.
هنگامی که محرک خارجی گسسته و مقدار آن بالاتر از یک حد آستانه معین باشد، پروتئینهایی که قبلاً ساخته شدهاند، از حوضچههای درون سلولی به محلی که سلول به طور موضعی به محرک خارجی، پاسخ میدهد حرکت کرده و ترجمه موضعی مورد بحث هم احتمالاً مکمل این حرکت خواهد بود. برای انجام این کار لازم است توزیع و شدت موضعی محرکهای خارجی موجود در محیطهای نانوالیافی سه بعدی به دقت و در ابعاد زیرسلولی (میکرون یا نانومقیاس) مهندسی شوند. این کار خصوصاً با توجه به آنکه در سالهای آینده دانشمندان به اطلاعات کمی و دقیقی برای آزمایش HTL دست مییابند، ضرورت بیشتری مییابد. به عنوان مثال، ویژگیهای نانومقیاس روی زیرلایههای مهندسی شده، (از قبیل لیگاندهایی که الگودهی آنها به جای یک لایه یکنواخت، به طور گسسته است) میتواند نشاندهنده پاسخهای شدیدتر سلول در نقاط تحریک شده باشد.
● ساخت محیطهای نانوالیافی سه بعدی
در حال حاضر ساخت نانوالیاف عمدتاً به دو روش الکتروریسی و پپتیدهای خودآرا (SAP) انجام میشود اما در اینجا به برخی دیگر از فناوریها در این زمینه که بر اساس جداسازی ترکیبات پلیالکترولیت و جداسازی فازی انجام میشود هم اشاره میشود. در اکثر این روشها که در مهندسی بافت کاربرد دارد سعی میشود، بیشتر و بهتر با ECM طبیعی عمل شود. همچنین قابلیت کنترل دقیق خواص مکانیکی و شیمیایی در این روشها، امکان بررسی سیستماتیک و کمی پاسخهای سلولی به محیطهای سه بعدی نانوالیافی را با دقتی در حد زیر سلولی فراهم میسازد و این مسئله به سهم خود ما را از طرح مواد زیستی آگاه میکند.
● ریسندگی الکتریکی
ریسندگی الکتریکی مواد پلیمری به دلیل تنوع زیادی که دارد به عنوان یک ابزار راحت برای ساخت داربستهای نانوالیافی پدید آمده است [۴۹ و ۴۸]. در این روش از ولتاژهای بالا (۲۰-۵ کیلو ولت) برای باردار کردن محلولهای پلیمری- که از یک نازل خارج میشوند، سپس به سمت یک سر بسترا كه روی زمینه جمع شده است شتاب داده میشوند- استفاده میشود. در این روش حلال موجود در جِت محلول پلیمری تبخیر شده و در نتیجه تودهای بافته نشده از نانوالیاف پلیمری روی این سربسترا جمع و روی زمینه تشکیل میشود [۵۰]. خواص نانوالیاف به دست آمده از این روش (از قبیل قطر نانو رشته) را میتوان به آسانی و از طریق دستکاری محلول پلیمری و پارامترهای عملی مربوط به ریسندگی الکتریکی کنترل نمود [۴۸ و۵۰ و ۵۱].ساخت مدلهای سه بعدی ECM برای ایجاد ارتباط کمی بین پاسخهای سلولی نسبت به محیطهای نانوالیافی سه بعدی کاملاً تعریف شده پیرامون سلول و نیز استفاده در مهندسی بافت انجام میشود. این نانورشتهها را میتوان با ریسندگی الکتریکی پلیمرهای مصنوعی و یا مواد زیستی طبیعی به دست آورد. و سپس به منظور افزایش زیستسازگاری و کارکرد سلولی، آنها را با موتیفهای شیمیایی وظیفهدار كرد (functionalized] (۵۳]. داربستهای نانوالیافی به دست آمده در این روش، با موفقیت در کاشت انواع مختلفی از سلولهای مورد استفاده در مهندسی بافت (از قبیل سلولهای غضروف، استخوان، رگهای خونی شریانی یا سرخرگها، بافت قلب و تاندونها) بهکار برده شده است.
همچنین از دنبال هم قرار دادن این نانورشتهها میتوان مسیری را ایجاد نمود که سلولها در آن راستا قرار گیرند [۵۴ و۵۵]. به طور كلی نانوفیبرهای تولید شده به روش ریسندگی الكتریكی یك ماده پوششی قوی (mat) صاف ایجاد میكنند كه به صورت سه بعدی محدود شدهاند و به علاوه اندازه کوچک حفره این شبکهها، باعث برون دقت (infiltero tion) سلولها میشود. این مشکلات را میتوان با ریسندگی الکتریکی این نانوالیاف روی داربستهای میکروالیافی و استفاده از نازلهای چندگانه برای رسوبدهی لایههای نانورشتهای [۵۸] یا ریسیدن توأم سلول و رشتهها، برطرف نمود [۵۹].
● پپتیدهای خودآرا
روش دیگر طراحی و مهندسی نانورشتهها، بر اساس آرایشبندی الیگوپپتیدهای۱ مصنوعی به منظور ایجاد شبکهای الیافی برای حفظ وضعیت سه بعدی سلولها است. پپتیدهای مصنوعی، بلوکهای ساختمانی ایده آلی برای ساخت داربستهای نانوالیافی سه بعدی به شمار میآیند چرا که اولاً زیست سازگار بوده و ثانیاً میتوان آنها را طوری طراحی نمود که موتیفهایی از قبیل پپتیدهای RGP (مشابه آنچه ECMهای طبیعی یافت میشود) در تنظیم فتوتیپهای سلولی، نقش داشته باشد [۶۰ و ۶۱]. علاوه بر اینها به مرور زمان و با افزایش اطلاعات دانشمندان از پیچخوردگی پروتئین و برهمکنشهای پروتئین- پروتئین، طراحی الیگوپپتیدهای خودآرا آسانتر شده است [۶۲]. با تحریک این SAPها شبکهای از نانوالیاف سه بعدی در شرایط فیزیولوژیکی پیرامون سلولها شکل میگیرد و به این ترتیب یکی از مشکلات اصلی مربوط به استفاده از داربستهای نانوالیافی پیش ساخته برطرف میشود.
SAPها را میتوان از اسیدهای آمینه طبیعی [۶۲ و ۶۰و ۹و ۸] یا مصنوعی [۶۳] ساخت؛ اگر چه که حالت اول معمولاً بیشتر از حالت دوم در همانندسازی موتیفهای پروتئینی طبیعی (که با سلولها برهمکنش میکنند) کاربرد دارد. SAPهای محیط نانوالیافی خارجی پیرامون سلول بر اساس ترکیبات مارپیچی یا ورقهای در سطح ساختار ثانویه شکل میگیرند. این مواد به دلیل ضعف استحکام مکانیکی غالباً مستقیماً به صورت وسایل حمل سلولی یا دارورسانی به بیمار داده میشوند [۶۴] یا در داربستهای میکرورشتهای برای مهندسی بافت و کاربردهای سلولی مورد استفاده قرار میگیرند .
محققان پیشبینی میكنند که در آینده "چارچوب طراح" با میکرو محیطهای تنظیم کننده دقیقی برای نگه داشتن انواع مشخصی از سلولها به وجود آید [۶۶]. این داربستها به ویژه در تعیین پاسخهای سلول به محیطهای نانوالیاف سه بعدی جالب توجه هستند. به عنوان مثال داربستهای SAP میتوانند میکرو محیطهای ساخت رگ که سلولهای آندوتلیومی عروق کوچک در آن ساخته میشود را ۲ بهتر از کلاژن شبیهسازی كنند و در نتیجه مدل خوبی برای درک فرآیند رگزایی ۳ ساختارهای ایجاد شده با مهندسی بافت فراهم نمایند [۶۷]. فرآیند رگسازی برای وادار کردن سلولهای تولید رگهای خونی در غذارسانی به ساختارهای بزرگ ایجاد شده با مهندسی بافت، ضروری است و یکپارچه شدن این ساختار با بافت میزبانی که در آن کاشته میشود، را تسهیل میكند.
● جداسازی ترکیبات کمپلکس پلیالکترولیت
یک محیط نانوالیافی را میتوان با جداسازی ترکیبات الکترولیتهای با بار مخالف- روشی که در ابتدا برای ماکرو یا میکرو کپسوله کردن سلولها بهکار میرفت- نیز ساخت [۷۰-۶۸]. استفاده از این روش در شرایط مربوط به محیطهای آبی ملایم هم امکانپذیر است و بنابراین میتوان آن را در شکلگیری اولیه نانورشتهها و در خود in situ سلول به کار برد.
دانشمندان قبلاً تعداد زیادی از این پلیالکترولیتها را برای کپسوله کردن سلول توسعه داده و ارزیابی نموده بودند [ ۶۹ و ۶۸]. نانوالیاف کمپلکس جدا شده به این روش معمولاً استحکام مکانیکی ندارند و بنابر این قرار دادن آنها در چارچوبهای سه بعدی که در مهندسی بافت به کار میروند، دشوار است. این نانوالیاف را به آسانی میتوان در ساختارهای کشت سلولی از قبیل داربستهای میکروالیافی سه بعدی و کانالهای میکروسیالی که برای تثبیت و نگهداری سلولهای حساس و وابسته به تکیهگاه به کار میروند، مورد استفاده قرار داد [۷۲ و ۷۱]. در روش جایگزین، نانورشتهها ممکن است مستقیماً در میکرورشتهها قرار داده شده و از ویژگی نانورشتهای آنها برای نگهداری نمونههای کشت شده انواع مختلفی از سلولهای پستانداران استفاده شوند [۷۵ و ۷۲].
● جداسازی فاز
از روش جداسازی فاز- که معمولاً برای ساخت داربستهای میکروالیافی بهکار میرود- میتوان در ساخت داربستهای نانوکامپوزیتی نیز استفاده كرد. نواحی پلیمری و غنی از حلال یک محلول پلیمری را میتوان یا با سرد کردن محلول و یا با تعویض غیرحلال با حلال از هم جدا نمود. با استفاده از روش القایی جداسازی فاز گرمایی و با پلی(ال – لاتیك اسید)، یک داربست نانوالیافی سه بعدی (رشتههایی به قطر ۵۰۰-۵۰ نانومتر) ساخته میشود [۷۶]. این داربست دارای نسبت سطح به حجم و تخلخل قابل کنترل بالا (تا ۵/۹۸ درصد) است و خواص مکانیکی تعریف شدهای هم دارد. اخیراً دانشمندان به روشی دست یافتهاند که با استفاده از آن میتوان داربستهایی با شکل و اندازه دلخواه ساخت [۷۷].
در این روش که SFF (ساخت معکوس شکل آزاد جامد) نامیده میشود، میتوان بافت داربست و ابعاد آن را با یکپارچه کردن گویهای پارافینی برای اتصال حفرهها کنترل كرد. به این ترتیب امکان ساخت داربستهای با شکل و اندازه دلخواه فراهم میشود. توزیع نانوالیاف و یکنواختی آنها به قابل کنترل بودن فرآوری آنها بستگی دارد. ساخت این داربست که طی یک فرایند پنج مرحلهای انجام میشود، تاکنون تنها در پلیمرهای معدودی چون پلی (L- لاكتیك اسید) و ترکیبات آن بهکار رفته است [۴۹].
● تعیین مشخصات محیط نانوالیافی سه بعدی
با توجه به پیشرفتهای حاصل شده درروشهای میکروسکوپی و پروبهای فلورسانس، امکان تصویربرداری دینامیک از شبکههای نانوالیافی سه بعدی و برهمکنش آن با سلولها فراهم شده است. معمولاً پروپهای فلورسانس با تغییر دادن خواص ماتریکس یا برهمکنش با سلولهای یک سیستم بیولوژیکی میتوانند تا حد معینی در آن تداخل ایجاد كنند. دانشمندان در حال حاضر سعی زیادی دارند تا بتوانند رنگهای فلورسانس با خاصیت تهاجمی کمتر و پروتئینهایی اختصاصیتر ایجاد كنند و از آن برای مشخص كردن خواص شیمیایی محیط سلولی استفاده نمایند. برای کمی نمودن ساختار دینامیک ساختاری ECM بدون استفاده از پروپهای فلورسانس میتوان از روشهای تصویربرداری مختلف و روشهای حسگری نیرو از قبیل میکروسکوپ نیروی اتمی (AFM) و انبرکهای نوری استفاده نمود که قابلیت بالایی برای اندازهگیری خواص مکانیکی دارند.
● تعیین مشخصات ساختاری با استفاده از روشهای تصویربرداری
محققان برای مشاهده برهمکنشهای سلولی با شبکههای نانوالیافی سه بعدی از میکروسکوپ DIC استفاده میكنند که نوعی روش تصویربرداری افزایش دهنده وضوح به شمار میآید [۱۰۵]. از آنجا كه تصاویر دوبعدی به دست آمده از این روش به عمقهای متفاوت مربوط میشوند، به لحاظ ذاتی ایجاد تصاویر سه بعدی بر اساس تصویرهای DIC حتی پس از انجام فرآیند انطباق تصاویر هم دشوار است. در میکروسکوپ بازتابی یا برگشت پراش هم کانون از خواص مربوط به پراش نور از مولکولهای نانوالیافی ECM برای ایجاد تصویر استفاده میشود. از این روش میتوان برای تصویربرداری از رشتههای کلاژنی و برهمکنش سلول- ECM استفاده نمود [۱۰۶، ۵]. شکل (۶ ) تصویری از یک شبکه کلاژنی سه بعدی است که با استفاده از میکروسکوپ هم کانون برگشت پراش نوری با طول موج ۴۸۸ نانومتر به دست آمده است.در تصویربرداری سه بعدی به دلیل ایجاد پراش نور از عمق کانونی بیش از۳۰ ،کیفیت و میزان تفکیک تصویر بسیار بد است. دانشمندان برای رفع این مشکل از تحریک چند طول موجی استفاده میکنند تا به این ترتیب به تفکیک و عمق نفوذ مناسب دست یابند [ ۱۰۷]. برای افزایش وضوح تصویرها میتوان از تزئینات فلزی از قبیل نانوذرات طلا در نمونه استفاده نمود [۱۰۸]. SHG (تولید هماهنگ مرحله دوم) یک فرآیند نوری مرتبه دوم غیرخطی است که طی آن دو فوتون به طور به هم پیوسته، به صورت یک فوتون واحد با انرژی دوبرابر پراکنده میشوند. SHG برخلاف فلورسانس یک روش غیرجذبی است و هیچ آسیب فوتوشیمیاییای به نمونه نمیرسد.
برای اولین بار از SHG در یک میکروسکوپ نوری برای مشاهده ساختارهای میکروسکوپیک بلور استفاده شد [۱۰۹]. همچنین دانشمندان توانستهاند با استفاده از این روش تصاویری از ساختارهای کلاژنی تاندوم دم موش [۱۱۰]، پوست [۱۱۳-۱۱۱]، قرنیه [۱۱۵ و ۱۱۴]، مغز [۱۱۶]، تومورها [۱۱۷]، جنین ماهی [۱۱۸] به دست آورند. با افزایش حساسیت و تفکیک این میکروسکوپ هماکنون برای تصویربرداری از نانورشتههای سه بعدی ECM حتی در بافتهایی با پراش نوری قابل ملاحظه (چون بافت كبد) هم از این میكروسكوپ استفاده میشود. در اینجا سیگنال SHG بدون هر گونه نشانهگذاری خارجی( برچسبزنی بیرونی) توسط نانورشتههای کلاژنی تولید میشود. از آنجا كه استفاده از نورمادون قرمز و تابع کاهش یافته پخش نقطهای مؤثر تا حد زیادی باعث از بین رفتن تأثیر مخرب پراش نور میشود [۱۱۹]، عمق تصویربرداری در این روش حداقل صدها میکرون افزایش یافته و حتی برای نمونههای شفاف به بیش از یك میلیمتر رسیده و تفکیک و کیفیت تصویر هم بهبود مییابد.
نانوالیافهای سه بعدی موجود در محیط خارج سلولی و از جمله رشتههای کلاژنی، غالباً به لحاظ نوری غیرهمگن هستند و لذا میتوان آنها را با توجه به انكسار دوتایی میکروسکوپ پلاریزان آشکار نمود. در این حالت نمونه را تحت تابش نور قطبی شده خطی قرار داده و سپس این نور را توسط یک پلاریزور عمودی تجزیه میکنند. انكسار دوتایی نمونه، هم به شدت نور تبدیل شده و توسط یک آشکارساز ثبت میشود. میکروسکوپ پلاریزان کاربرد وسیعی در تعیین مشخصات کمی کلاژن در بافتهای رنگآمیزی نشده دارد [۱۲۰]. معمولاً به منظور افزایش انكسار، نانورشتههای کلوژنی بافتهای ثابت را با برچسبی از نوعی رنگ قرمز مشخص میکنند و به این ترتیب مولکولهای رنگ در راستای رشتههای کلاژنی قرار میگیرند [۱۲۱]
● تعیین مشخصات شیمیایی با استفاده از میکروسکوپ همکانون فلورسانی
برچسبزنی (به عنوان مثال با استفاده از پروتئینهای فلورسانس)، یکی دیگر از روشهای پویای تصویربرداری از داربستهای نانورشتهای سه بعدی به شمار میآید [۱۲۲]. به طور کلی، تصویربرداری به کمک برچسبزنی فلورسانس، اختصاص بیولوژیکی بالا و وضوح خوبی دارد اما در مقابل باعث ایجاد تداخل در یکپارچگی ساختاری محیط نانوالیافی و نیز پاسخهای سلولی میشود. دانشمندان برای از بین بردن اثرات تداخلی نامطلوب پروتئینهای فلورسنت بزرگ، روشهای برچسبزنی مولکول کوچک مورد توافق promising را ابداع نمودهاند [۱۲۳]. انتقال انرژی تشدید فلورسانس (FRET) یک خطکش نوری است که میتوان با استفاده از آن فاصله بین فلوروفورهای دهنده و گیرنده را اندازهگیری نمود. این کار با مشخص نمودن جفتشدگی میدان نزدیک از طریق تعیین شدت فلورسانس یا طول عمر آن انجام میشود.
با استفاده از این روش (که امکان تمایز بین ساختارهای مختلف پروتئینی را فراهم میکند) میتوان پی برد که چگونه سلولها ساختار پروتئینی را در محیط بیرونی خود تغییر میدهند و دریافت که چگونه این تغییرات ساختاری با تغییر کارکرد سلول در ارتباطند [۱۲۵ و ۱۲۴]. دانشمندان با نشاندار كردن (tagged) فیبرونکتین به فلورفورهای دهنده و گیرنده در نقاط مختلف و FRET بین مولکولی مشاهده كردند که فیبرونکتین در پاسخ به کشش اسكلت (داربست) سلولی ایجاد شده توسط فیبروبلاستهای NIH۳T۳، دچار فرورفتگی شده و کشیده میشود. همچنین فلورسانس حاصل از تحریک دوفوتونی هم موجب افزایش عمق تصویربرداری به صدها میکرون شده و امکان مشاهده ساختمان بافت در عمق هم فراهم میشود [۱۲۶].
● تعیین مشخصات مکانیکی با استفاده از روشهای حسگری نیرو
با توسعه روشهای حسگری نیرو، امکان اندازهگیری نیروهای ضعیف تا حد پیکونیوتن و یا حتی کمتر از آن هم فراهم شده است. به علاوه اندازهگیریهای مربوط به شکل هندسی و خواص مکانیکی (از قبیل ثابت فنر و مدول یانگ) تکتک نانورشتهها را هم میتوان با این روشها به دست آورد. میکروسکوپ نیروی اتمی (AFM) حدود بیست سال قبل به عنوان یک روش تصویربرداری با قدرت تفکیک بالا اختراع شد [۱۲۷] با این میکروسکوپ علاوه بر اندازهگیریهای نقشهبرداری، میتوان نیروهای در حد پیکونیوتن و جابجاییهای نانومتری را هم آشکار کرد و به این ترتیب بر اساس منحنیهای نیرو- فاصله مطالعاتی را در زمینه نیروهای درونی و بین ملکولی انجام داد [۱۲۸].
محققان با استفاده مؤثر از AFM و اندازهگیری نیروی پیوندی اینتگرین و مولکولهای های ECM [۱۳۱ و ۱۲۹] و ارتباط دادن آن با استحكام نانورشتههای طبیعی [۱۳۲] و مهندسی شده [۱۳۳]، میتوانند به بررسی برهمکنشهای سلول- ECM بپردازند. همچنین با تلفیق AFM و میکروسکوپ نوری، یعنی قرار دادن نوک پیمایشگر آن روی یک میکروسکوپ نوری معکوس، میتوان خواص مکانیکی و اطلاعات ساختاری یا عاملی را به طور همزمان به دست آورد و ثبت نمود. به این ترتیب با تلفیق این دو مجموعه اطلاعات، میتوان به طور کمی از پاسخ سلولها به محیط نانوالیافی سه بعدی آگاه شده و به محدودیتهای مکانی- زمانی محرکهای شیمیایی و مکانیکی خارج از سلول هم پیبرد. با استفاده از انبرکهای نوری میتوان اجسام میکروسکوپی و حتی اتمهای منفرد را در دماهای پائین دستکاری نمود [۱۳۴ و ۱۳۵]. مبنای اساسی این روش فشار تابشی نور بر اجسام دیالکتریک است. نیروی به داماندازی یک اتم یا مولکول منفرد بسیار کم است، لذا برای دستکاری یک مولکول منفرد در دمای اتاق، محققان معمولاً دستههایی- به عنوان مثال دانههای پلیاستایرن پلیاسترین- را به آن متصل میکنند. انبرکهای نوری علاوه بر دستکاری اجسام ریز، در اندازهگیری نیروهای بینمولکولی در حد پیکونیوتن [۱۳۶] و تعیین کشسانی رشتهها، کاربرد دارند [۱۳۷]. با متصل نمودن دانههای پلیاستایرن به نانورشتهها، میتوان آنها را با استفاده از این انبرکهای نوری خم نموده و استحكام آنها را اندازه گرفت. همچنین انجام اندازهگیریهای سه بعدی در این روش بسیار آسانتر از به کاربردن AFM است.
انبرکهای نوری، علاوه بر تعیین مشخصه رشتههای منفرد، میتوانند مشخصه مربوط به خواص جریانشناسی حجمهای معین را هم از طریق اندازهگیری موضعی گرانروی کشسان تعیین نمایند [۱۳۸]. این خواص رئولوژیكی، به ساختار ماكرومولکولی یا فرامولکولیECM یا محیط نانوالیافی سه بعدی بستگی دارد.
● چشمانداز آینده
علیرغم وجود فناوریهای مهندسی و ساخت نانورشتههایی با ویژگیهای پیچیده، مطالعات سیستماتیک در سطح مولکولی درباره پاسخهای سلولی به شبکههای نانوالیافی سهبعدی، عمدتاً با ECMهای طبیعی انجام میشده است. در حال حاضر برای درک سیستماتیک برهمکنشهای سلول- شبکه و فرایند بازگشتی، طراحی مواد زیستی جدید با کاربردهای خاص، استفاده از چنین زیرلایههای نانوالیافی مهندسی شدهای، بسیار ضروت دارد. محصولات تجاری از این دست و زیرلایههای نانورشتهای بزرگتر موجب تسریع استفاده از چنین محیطهای سه بعدی در تحقیقات زیستشناسی سلولی و نیز مهندسی بافت میگردد.
با توسعه ابزارها و فرآیندهای جدید، داربستها و نانورشتههای سه بعدی با یکنواختی بیشتر و خواص مکانیکی/ شیمیایی قابل کنترل پدید خواهند آمد. تمامی این پیشرفتها، به همراه روشهای تصویربرداری و حسگری نیرو که با گذر زمان برای بررسی سه بعدی نانورشتهها پدید میآید، پشتوانه تحقیقات با دقتهای زیرسلولی درباره چگونگی پاسخ فرآیندهای سلولی و مولکولی مختلف به این داربستها را تشکیل میدهند. به این ترتیب، دانشمندان میتوانند LTH را عملاً مورد آزمایش قرار داده و آن را اصلاح نمایند. همچنین اندازهگیریهای سیستماتیک و کمی پارامترهای مولکولی و سلولی در نقاطی مجزا از برهمکنش سلول ECM و نیز در کل سطح سلول، قابلیت جدیدی را فراهم میکند که عبارتست از:
▪ امکان ایجاد مدلهایی محاسباتی و نیز مدلهایی از سیستمهای زیستشناسی که نشاندهنده پاسخهای سلولی نسبت به نانوالیافی است که دارای خواص مکانیکی و شیمیایی مختلف بوده و به طور زمانی و مکانی توزیع شدهاند. چنین مدلهایی به توسعه کاربردی بیشتر انواع جدیدی از محیطهای نانوالیافی منجر میشود.
منبع : ستاد ویژه توسعه فناوری نانو

















































































ایران مسعود پزشکیان دولت چهاردهم پزشکیان مجلس شورای اسلامی محمدرضا عارف دولت مجلس کابینه دولت چهاردهم اسماعیل هنیه کابینه پزشکیان محمدجواد ظریف
پیاده روی اربعین تهران عراق پلیس تصادف هواشناسی شهرداری تهران سرقت بازنشستگان قتل آموزش و پرورش دستگیری
ایران خودرو خودرو وام قیمت طلا قیمت دلار قیمت خودرو بانک مرکزی برق بازار خودرو بورس بازار سرمایه قیمت سکه
میراث فرهنگی میدان آزادی سینما رهبر انقلاب بیتا فرهی وزارت فرهنگ و ارشاد اسلامی سینمای ایران تلویزیون کتاب تئاتر موسیقی
وزارت علوم تحقیقات و فناوری آزمون
رژیم صهیونیستی غزه روسیه حماس آمریکا فلسطین جنگ غزه اوکراین حزب الله لبنان دونالد ترامپ طوفان الاقصی ترکیه
پرسپولیس فوتبال ذوب آهن لیگ برتر استقلال لیگ برتر ایران المپیک المپیک 2024 پاریس رئال مادرید لیگ برتر فوتبال ایران مهدی تاج باشگاه پرسپولیس
هوش مصنوعی فناوری سامسونگ ایلان ماسک گوگل تلگرام گوشی ستار هاشمی مریخ روزنامه
فشار خون آلزایمر رژیم غذایی مغز دیابت چاقی افسردگی سلامت پوست