یکشنبه, ۱۶ اردیبهشت, ۱۴۰۳ / 5 May, 2024
پاسخ های سلولی به محیط نانوالیافی
سلولها دربرابر سختی و انعطافناپذیری مکانیکی، نانوتوپولوژی سه بعدی زیرلایهها و همچنین محدودیت زمانی و مکانی محرکهای برون سلولی، پاسخ شدیدی میدهند. در این مقاله خلاصهای از آخرین پیشرفتها در زمینه کشف و طراحی محیطهای زیستسازگار نانوالیافی برونسلولی در سطح مولکول و سلول (که در مهندسی بافت وتحقیقات زیستشناسی کاربرد دارد) ارائه میشود و به این ترتیب زمینه توسعه بیشتر داربستهای نانوالیافی سه بعدی به منظور تعیین پاسخ خاص سلول در این کاربردها فراهم میشود.
سلولها در محیط زنده بدن توسط یک ماتریکس شبکهای برونسلولی و سه بعدی از نانوالیاف ۱(ECM) دارای لیگاندهای شیمیایی متفاوت که با پذیرندههای سطح سلول برهمکنش میکنند، نگهداری میشوند [۴-۱]. آنچه باعث ارتباط ECM و سلول میشود فرایند دینامیکی و پیچیدهای است که طی آن خواص شیمیایی و فیزیکی ECM، موجب پاسخهای سلولی متفاوتی میشوند [۵ و۱]. بنابر این برای آنکه یک بستر مهندسی شده دقیقاً همان کار ECM طبیعی را انجام دهد، باید همان (نقشه) نانوالیافی و سه بعدی بودن و نیز فراوانی موتیفهای شیمیایی آن را دربرداشته باشد [۷و۶]. پیشرفت فناورینانو در سالهای اخیر امکان طراحی و مهندسی مواد زیستی جدید با این سطح از پیچیدگی را فراهم کرده است [۹-۷].
استفاده از این مواد زیستی پیچیده برای کاربردهای خاص توسط مصرف کننده نهایی، مستلزم انجام یک فرایند محاسباتی بازگشتی برای درک مکانیسم هدایت کننده برهمکنشهای سلول- شبکه میباشد، تا به این ترتیب بتوان خواص ماده زیستی را کنترل کرد و در نهایت به پاسخهای سلولی مناسب و دلخواه دست یافت . در اینجا ما بر اهمیت ویژگیهای نانوالیاف سه بعدی محیط برونسلولی و نقش آن در تنظیم پاسخهای سلولی دارای تفکیک موضعی یا زیرسلولی به روشی وابسته به زمان- مکان تأکید میکنیم. همچنین فناوریهای نوین ساخت و تعیین مشخصات محیطهای نانوالیافی در کاربردهای مرتبط با آن، از دیگر مواردی است که در این مقاله بیشتر به آن توجه میشود.
● تأثیر محیط نانوالیافی بر پیامرسانی و فنوتیپ۱ سلول
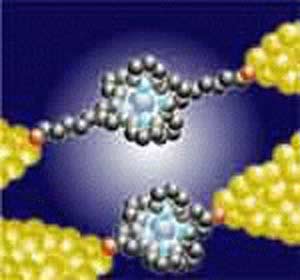
سلولها از طریق پروتئینهای سطحی خود از قبیل اینتگرینها۲ با محیط نانوالیافی خارجی پیرامون خود برهمکنش میکنند و در اثر این برهمکنشها، مسیرهای مختلف سیگنالدهی که فرایندهای سلولی از قبیل شکل سلول، تحرکپذیری و تكثیر سلولی را تنظیم میکند، فعال میشوند (شکل ۲). آگاهی از این پاسخهای (زیستی) ویژه، که با جنبههای مختلفی از محیط نانوالیافی تحریک میشوند- در هدایت طرحها و مهندسی بسترهای جدید که به تقلید از برهمکنشهای سلول- ماتریکس داخل بدن، ساخته میشود اهمیت بسزایی دارد.
● تأثیر توپوگرافی سه بعدی
برخلاف بسترهای دوبعدی پهن و صلب، این نانوالیاف سه بعدی امکان برهمکنش سلولها با یک حلقه سهبعدی انعطافپذیر را فراهم میکند. این نانوالیاف سه بعدی همچنین باعث ایجاد فتوتیپهای سلولی مشابه فتوتیپهای درون بدن میشود و سلولی شکلزایی بافت را نیز افزایش میدهند [۱۳-۱۰]. رویدادهای مولکولی مربوط به برهمکنش سلولها با این نانوالیاف سهبعدی نیز متفاوت از موارد مشابه در بسترهای دوبعدی از قبیل ظرفهای کوچک مصنوعی کشت میکروب میباشد. سه بعدی بودن این نانوالیاف به تنهایی در فعال کردن مسیر تبدیل سیگنال با واسطه Rac۳ داخل فیبرو بلاستهایNIH۳T۳ و سلولهای عادی کلیه موش، نقش دارد، به خصوص اگر روی نانوالیاف الکتروریسی۴ شده و در غیاب ماکرومولکولهای ECM کشت شوند [۱۴].
حرکت فیبروپلاست در شبکههای کلوژنی سه بعدی (که در شکلگیری دوباره کلوژن طی روند بهبودی زخم اهمیت دارد) ، تا حد بسیار زیادی به زنجیر سنگین میوزین غیرعضلانی Nanomuscle Myosine Heavy Chain II-B۱ بستگی دارد. این در حالی است که حرکت سلول روی سطوح دو بعدی صلب از این ویژگی برخوردار نیست [۱۵]. پاکسیلین (در چسبندگی موضعی) و اینتگرین (در چسبندگی رشتهای) ، برخلاف بستر دوبعدی با دو ناحیه چسبندگی مجزای موضعی و رشتهای کلاسیکی روی آن، حالت سه بعدی موضعی شده مشترکی پیدا میکنند. [۱۶]. اعمال فشار مکانیکی و پهن کردن مصنوعی ماتریکسهای نانوالیافی سه بعدی به منظور ایجاد شبکههای دو بعدی صاف مجازی، باعث از دست رفتن مشخصه موضعیسازی مشترک سهگانه اینتگرین، پاکسیلین و فیبرونکتین موجود در چسنبدگیهای شبکه سهبعدی میشود [۱۶]. لذا نقشهبرداری نانوالیافی سه بعدی، تأثیر بسیار زیادی بر برهمکنشهای سلول-ECM در سطح مولکولی خواهد داشت.
● تأثیر خواص مکانیکی
سلولها به لیگاندهای شیمیایی و نیز محرکهای مکانیکی متنوع محیط پیرامونی خارجی پاسخ میدهند [۱]. پخش کردن سلولها روی بسترهای صلب باعث تکثیر آنها میشود و lineage differenrition تمایز دودمان و سلولهای بنیادی (stemcell) را تغییر میدهد [۱۹]. سلولها به نیروهای حاصل از تنش برشی و فشردگی حساس بوده و در نتیجه آن مدلسازی مجدد و خواص مکانیکی ساختار بافت تحت تأثیر قرار خواهد گرفت [۲۳-۲۰]. در وضعیت سه بعدی، باز هم سلولها به صلب بودن نانوالیاف حساس هستند [۲۵ و ۲۴و ۱۶]. نیروهای خارجی هم از طریق مسیر ارتباطی ECM- اینتگرین- سیتواسکلتون۲ آرایش چسبندگی موضعی۳ که تیروزین کیناز فسفاتاز و كینازهایSRC SFKS را تحریک کرده و موجب فعالسازی پروتئینهای G کوچک و کینازهای MAP۴ و در نهایت تنظیم رویدادهای پایین دستی سلولی میشوند [۲۶]. باید توجه داشت که کمی کردن سیستماتیک این رویدادها و نیز ایجاد مدلهایی از پاسخ سلول به خواص مکانیکی و شیمیایی نانوالیاف به منظور هدایت روند توسعه محیطهای نانوالیافی سه بعدی به سمت مصرف و کاربرد، اهمیت دارد.
● تأثیر محدودیتهای زمانی- مکانی بر محرکهای برونسلولی
پاسخهای سلولی به خواص فیزیکی و شیمیایی نانوالیاف سه بعدی، غالباً به مکان و زمان بستگی دارد [۲۹-۲۷]. هنگامی که سلولهای بنیادی عصبی و سلولهای ماهیچهای صاف۵ به روش الکتروریسندگی کشت شوند، خود را در امتداد این الیاف کشیده و جهت میدهند [۳۱ و ۳۰]. حساسیت نانوالیاف به ترتیب مکانی، احتمالاً ناشی از برهمکنشهای موضعی سلولها با محرکهای برونسلولی است که در سطح تفکیک زیرسلولی روی میدهند. اغلب پروتئینهای ساختاری یا سیگنال دهندهای که در حسگری صلب، (تکیهگاه ماتریکسی برون سلولی) و مهاجرت سلولی حضور دارند، در نقاط خاصی از سلول از قبیل نوک جلویی فیبرو بلاستهای مهاجر یا نقاط مربوط به چسبندگی موضعی روی میدهد و در نقاط دیگر و حتی در فاصله ۵/۰ میکرومتر دورتر، چنین پدیدهای روی نمیدهد [۲۸].
یک اتصالECM- اینتگرین- سیتواسکلتون منفرد میتواند در نقطه معینی از سلول و در پاسخ به نیرویی خارجی از سلول و بدون تحت تأثیر قرار دادن اتصال مجاور ایجاد شود[ ۳۲ و۳۳]. خوشه شدن مولکولهای اینتکرین و تشکیل چسبندگی موضعی و الیاف فشاری ، تنها هنگامی روی میدهد که تعداد کافی از جایگاههایRGD، داخل ناحیه مجزایی با اندازه کوچکتر از ۷۰ نانومتر جمع شوند [۲۹-۲۷].
همچنین مولکولهای همراه با اتصالات اینتگرین- سیتواسکلتون موجود در چسبندگی سلول- ECM، به سخت بودن این نانوالیاف ECM حساس هستند [۳۸-۳۵و ۳۲] و طی چرخه زندگی نقاط چسبندگی، همراه با گذشت زمان، تغییر میکنند [۳۴]. شکلگیری تماسهای سلولی با ECM یک فرایند پیوسته نیست اما شامل چرخههای انقباض و رها شدن است. دانشمندان اخیراً نشان دادهاند که وجود نانوالیاف خارجی درهپاتوسیتها (سلولهای کبدی) در زمانها و توالیهای مختلف از کشت ساندویچی، پاسخهای متفاوتی را نشان میدهد [۳۹].
پس میتوان گفت سلولها نسبت به محدودیتهای زمانی و مکانی محرکهای محیط خارجی پیرامونی حساس هستند. به نظر میرسد مجموع پاسخهای موضعی سلول به این محرکهای برونسلولی طی زمان، بتواند به تعیین شکل و بیان ژنی سلول کمک نماید [۴۰ و ۲۶]. بنابراین مهندسی و درک پاسخهای موضعی سلولی به توزیع زمانی و مکانی محرکهای برون سلولی، در توسعه بیشتر محیطهای نانوالیافی سه بعدی پیرامونی اهمیت بسیاری دارد.
● تأثیر محرکهای برونسلولی بر پاسخهای موضعی سلولی
با توجه به تحریک حاصل از محرکهای برون سلولی (جدا از هم) و به حد کافی قوی، سلولها در ابتدا به طور موضعی و سپس همگی به این محرکها پاسخ میدهند. شبكه آندوپلاسمی ER۱ و بسیاری دیگر از ترکیبات مرتبط با شبكه آندوپلاسمی، معمولاً به صورت یکنواخت و با ساختاری شبکه مانند داخل سیستوپلاسم سلول و در یك محیط كشت دو بعدی توزیع میشوند . این ترکیبات در صورت قرار گرفتن در معرض تحریکات خارج از سلول، تماس سلولی، تکثیر و یا مهاجرت، میتوانند به صورت پویا آرایشی دوباره پیدا كنند.
هنگامی که دانههای پوشش شده با فیبرزنكتینبا سطح بالایی سلول برهمکنش میکند، برخی از پروتئینهای غشایی ER از قبیل کینکتین، پروتئین همراه با گیرنده(RAP) ، کالرتیکولین۱ [۴۱] و فاكتور طویل كننده یوركاریوتی((EEF-۱۹۴۶; ( EEF) به سرعت به محل ترکیبات اینتگریندار AC که به صورت موقت در اطراف بید پوشش شده با فیبرونكتین شکل گرفتهاند منتقل میشوند. این ترکیبات به فیبرونکتین پخش شده در محلول یا فیبرونکتینی که به طور یکنواخت روی یک سطح دوبعدی بزرگ پخش شده باشد، پاسخ نمیدهند (شکل ۴). پژوهش دانشمندان نشان داده شده است که EEF-۱۹۴۶; از طریق کینکتین باعث نگه داشتن کل ترکیب EEF روی غشاء ER شده و سنتز پروتئینهای غشاء/ ترشحی۲ و سیتوزولیک۳ را تنظیم میکند [۴۳].
به علاوه محققان مشاهده کردند که ریبوزومها و MRNA، IACهای اطراف دانههای پوشیده شده از فیبرونکتین را هدف گرفته و قبل از آنکه تغییرات نسخهبرداری را بتوان ردیابی نمود، سنتز پروتئینی سریعی روی میدهد [۴۵و ۴۱]. بنابر این میتوان سنتز موضعی پروتئینها را یک فرایند سلولی کلی فرض نمودکه در پاسخ به نشانهها و محرکهای قوی، گسسته و موضعی شده خارج از سلول به روشی وابسته به مکان و احتمالاً زمان روی میدهد (شکل ۵). بر اساس این فرضیه ترجمه محلی۴ (LTH)، کارخانه سنتز پروتئین یک سلول- شامل ریبوزومها، MRNA و تنظیم کنندههای انتقال (از قبیل میکرو RNA) به محلی نزدیک محرک خارجی که میتواند دانههای پوشیده شده با فیبرونکتین [۴۱]، نیروی وارد شده توسط انبرکهای نوری [۴۶] یا دیگر نقاط اتصال زیر لایه صلب باشند- حرکت میکند.
هنگامی که محرک خارجی گسسته و مقدار آن بالاتر از یک حد آستانه معین باشد، پروتئینهایی که قبلاً ساخته شدهاند، از حوضچههای درون سلولی به محلی که سلول به طور موضعی به محرک خارجی، پاسخ میدهد حرکت کرده و ترجمه موضعی مورد بحث هم احتمالاً مکمل این حرکت خواهد بود. برای انجام این کار لازم است توزیع و شدت موضعی محرکهای خارجی موجود در محیطهای نانوالیافی سه بعدی به دقت و در ابعاد زیرسلولی (میکرون یا نانومقیاس) مهندسی شوند. این کار خصوصاً با توجه به آنکه در سالهای آینده دانشمندان به اطلاعات کمی و دقیقی برای آزمایش HTL دست مییابند، ضرورت بیشتری مییابد. به عنوان مثال، ویژگیهای نانومقیاس روی زیرلایههای مهندسی شده، (از قبیل لیگاندهایی که الگودهی آنها به جای یک لایه یکنواخت، به طور گسسته است) میتواند نشاندهنده پاسخهای شدیدتر سلول در نقاط تحریک شده باشد.
● ساخت محیطهای نانوالیافی سه بعدی
در حال حاضر ساخت نانوالیاف عمدتاً به دو روش الکتروریسی و پپتیدهای خودآرا (SAP) انجام میشود اما در اینجا به برخی دیگر از فناوریها در این زمینه که بر اساس جداسازی ترکیبات پلیالکترولیت و جداسازی فازی انجام میشود هم اشاره میشود. در اکثر این روشها که در مهندسی بافت کاربرد دارد سعی میشود، بیشتر و بهتر با ECM طبیعی عمل شود. همچنین قابلیت کنترل دقیق خواص مکانیکی و شیمیایی در این روشها، امکان بررسی سیستماتیک و کمی پاسخهای سلولی به محیطهای سه بعدی نانوالیافی را با دقتی در حد زیر سلولی فراهم میسازد و این مسئله به سهم خود ما را از طرح مواد زیستی آگاه میکند.
● ریسندگی الکتریکی
ریسندگی الکتریکی مواد پلیمری به دلیل تنوع زیادی که دارد به عنوان یک ابزار راحت برای ساخت داربستهای نانوالیافی پدید آمده است [۴۹ و ۴۸]. در این روش از ولتاژهای بالا (۲۰-۵ کیلو ولت) برای باردار کردن محلولهای پلیمری- که از یک نازل خارج میشوند، سپس به سمت یک سر بسترا كه روی زمینه جمع شده است شتاب داده میشوند- استفاده میشود. در این روش حلال موجود در جِت محلول پلیمری تبخیر شده و در نتیجه تودهای بافته نشده از نانوالیاف پلیمری روی این سربسترا جمع و روی زمینه تشکیل میشود [۵۰]. خواص نانوالیاف به دست آمده از این روش (از قبیل قطر نانو رشته) را میتوان به آسانی و از طریق دستکاری محلول پلیمری و پارامترهای عملی مربوط به ریسندگی الکتریکی کنترل نمود [۴۸ و۵۰ و ۵۱].




























































































































































نمایندگی زیمنس ایران فروش PLC S71200/300/400/1500 | درایو …
دریافت خدمات پرستاری در منزل
pameranian.com
پیچ و مهره پارس سهند
تعمیر جک پارکینگ
خرید بلیط هواپیما
امیرعبداللهیان حسین امیرعبداللهیان اوکراین دولت سیستان و بلوچستان انتخابات جنگ مجلس شورای اسلامی حجاب دولت سیزدهم افغانستان مجلس
سیل هواشناسی تهران شهرداری تهران بارندگی سازمان هواشناسی باران فضای مجازی یسنا آتش سوزی هلال احمر آموزش و پرورش
هوش مصنوعی خودرو دلار قیمت خودرو قیمت دلار مسکن قیمت طلا تورم بانک مرکزی بازار خودرو حقوق بازنشستگان ارز
مسعود اسکویی تلویزیون ایتالیا صدا و سیما دفاع مقدس حج مهران غفوریان موسیقی صداوسیما سریال سینمای ایران سازمان صدا و سیما
رژیم صهیونیستی فلسطین اسرائیل جنگ غزه حماس روسیه آمریکا انگلیس یمن نوار غزه ایالات متحده آمریکا جنگ اوکراین
فوتبال پرسپولیس استقلال سپاهان رئال مادرید لیگ برتر باشگاه استقلال بازی باشگاه پرسپولیس علی خطیر جواد نکونام بایرن مونیخ
خواب آیفون اینستاگرام دیابت اپل ناسا صاعقه تبلیغات موبایل گوگل
سلامت کبد چرب فشار خون گرما